The Architecture of Tension: Piezo1 and Apical-Basal Remodeling Govern Epithelial Columnarization
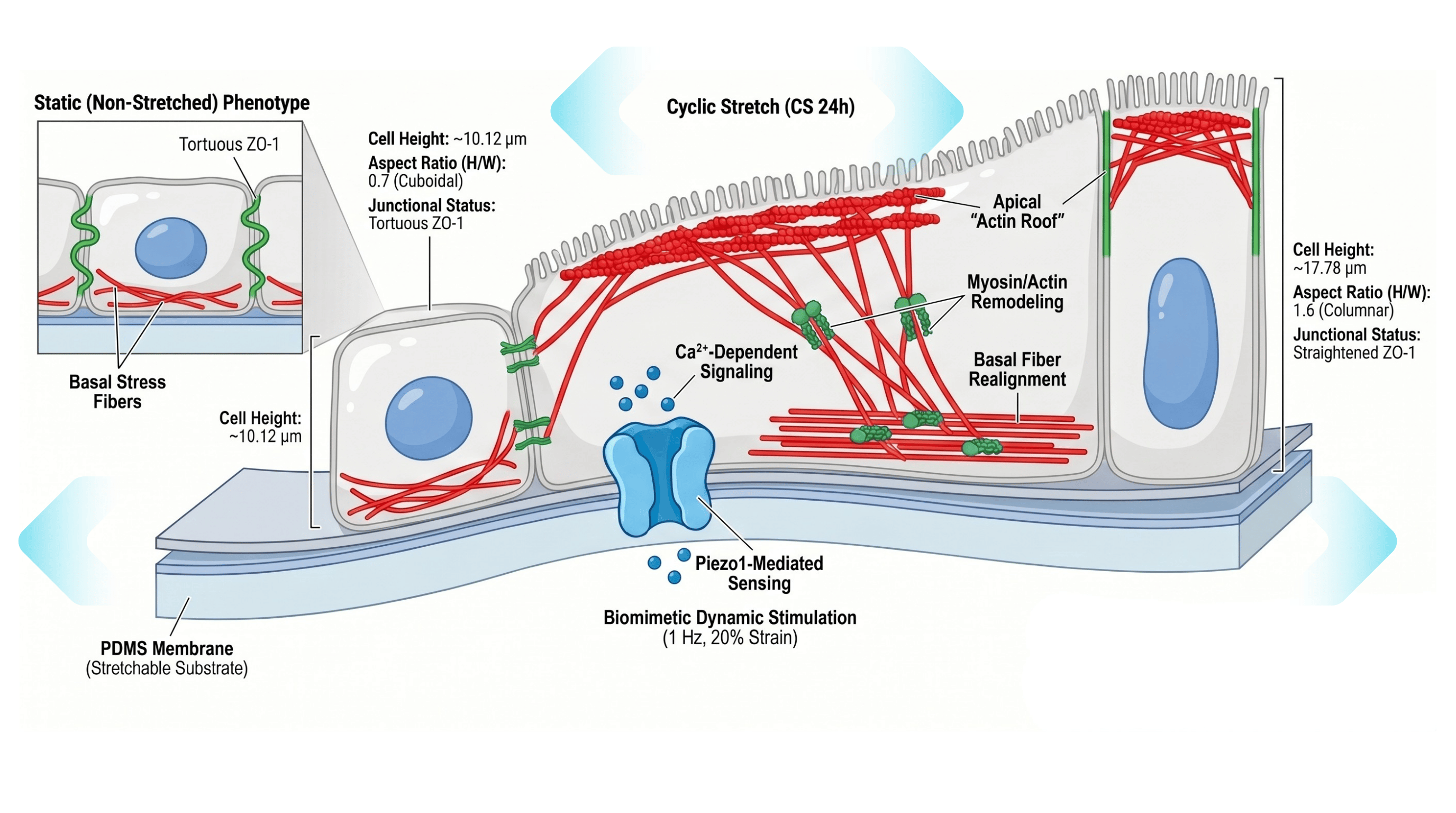
Epithelial shape is traditionally viewed as a static derivative of spatial confinement, yet living tissues exist within a continuous spectrum of physical deformation. This research challenges passive morphological paradigms by deploying a precise kinetic microenvironment: a 20% uniaxial cyclic stretch at 1 Hz sustained over 24 hours. These specific parameters abandon the artifactual silence of standard tissue culture, successfully recreating the intense, persistent biomimetic dynamics experienced by cardiovascular and gastrointestinal networks under hypertensive or atherosclerotic strain. Through this controlled mechanical tension, the epithelium does not merely survive physical stress—it initiates a highly coordinated, active biomechanical awakening to preserve tissue homeostasis.
The Mechanotransduction Circuit: From Basel Strain to Apical Polarization
The conversion of physical pulling into architectural reorganization follows a strict spatial hierarchy. The cyclic strain initially engages the basal substrate, triggering the immediate activation of the mechanosensitive ion channel Piezo1 on the plasma membrane. Functioning as the primary molecular "promoter" of the mechanical cascade, Piezo1 translates physical tension into an intracellular biochemical flux, propagating a bidirectional basal-to-apical transduction network.
Upon intercepting this signal, the cellular machinery orchestrates a sophisticated spatial redistribution of mechanical force. At the apical domain, actin monomers rapidly polymerize to forge a protective, contractile "actin roof." Concurrently, at the basal layer, stress fibers realign perpendicularly to the vector of stretching—an active structural defense deployed to dissipate the external mechanical load.
Tight Junctions and the Mechanics of Columnarization
The final phase of this morphological metamorphosis relies on the amplification of intercellular tension. Under the influence of cyclic strain, the tight junction protein ZO-1 transitions from a relaxed, tortuous configuration into a taut, linearized state. This structural straightening recruits a dense population of non-muscle myosin IIB, drastically elevating the cell's apical-basal tension.
This intracellular pulling force effectively elongates the flat, cuboidal monolayer into a highly functional, columnar epithelial phenotype. Ultimately, epithelial columnarization transcends the definition of a passive compromise forced by crowding; it represents an active molecular recalibration driven by biomimetic dynamics.
A static substrate merely permits cells to subsist; only through pathologically calibrated pulsation do epithelial sheets repack their cytoskeleton and betray the hidden mechanics of tissue homeostasis.